Have you ever wondered why you get a fever when you're sick?
Mace et al. (2011) also wondered this- they hypothesized that cells that have been exposed to higher than normal temperatures (in the human body) would exhibit an increased immune response. If this were the case, it would be highly adaptive, or good for the human body to induce fever whenever it recognized a pathogen in its body.
Does fever make our immune system more effective?
The type of immune response that Mace et al. (2011) studied was production of cytotoxic lymphocytes, also known as T-cells, and their differentiation into effector cells (a specialized T-cell). At early stages, cells have to ability to differentiate, or change into specific types of cells as the body needs them (see: stem cells and medical research).
 |
| Figure 2. Cytotoxic T-Cells lyse foreign cells |
The Mace et al. 2011 study
In order to test if heat increased T-cell differentiation into effector cells, Mace et al. (2011) set up a simple experiment. They first exposed cells to three temperatures: 33 °C (= 91.4 °F), 37 °C (= 98.6 °F), and 39.5 °C (= 103.1 °F), which are below normal, average, and above normal temperatures for a human. Then, the researchers mixed in non-host material and counted the number of T-cells that differentiated into effector cells.
Again, Mace et al. 2011 hypothesized: T-cells exposed to higher than normal temperatures will experience increased differentiation into effector cells
The null hypothesis which they would compare against was: T-cells exposed to higher than normal temperatures will experience the same amount of differentiation into effector cells as the other temperatures
The result of Mace et al. 2011
The result that Mace et al. (2011) found (FYI- this isn't the only result, if you are interested, I encourage you to read the full study) was that there was increased differentiation into effector cells when the cells were first warmed to 39.5 °C, relative to the other two temperatures. Meaning, while fever may be uncomfortable, it can make your immune system more effective. It has been noted though- fevers higher than 39.5 °C (= 103.1 °F) can be very dangerous, and one should seek medical attention if they last for any period of time.
Wait- so how does this all relate to the insect immune system?
All organisms share a common ancestor (insects are no exception), and thus, there is considerable overlap in structure (how it's built) and function (what it's built for) of their bodies. Secondly, similar features that serve similar purposes tend to arise through the process of independent, or convergent evolution. In the case of the insect immune system, it has similar immune responses to non-host material as described above.
Insect internal anatomy
One of the main differences of the insect body, relative to that of a human is it has an open circulatory system. Humans have vessels and arteries that transport fluids and nutrients throughout the body. Insects on the other hand have a large body cavity known as a hemocoel. This can make movement of some fluids and nutrients tough for insects (although they have figured out plenty of ways to get around this), but easy for insect physiologists to study.
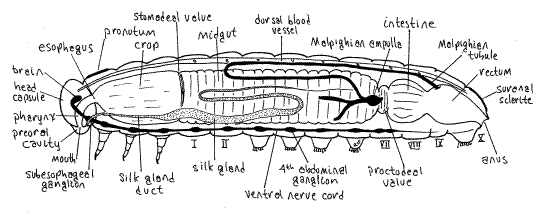 |
| Figure 3. Generalized internal anatomy of an immature lepidopteran |
I
Immune response in insects
 |
| Figure 4. The generalized morphology of some typical hemocytes that are found in insect hemolymph. From Woodring (1985). |
Insects have what is known as innate immunity (as described above). This is the ability to distinguish between self and non-self. Innate immunity in insects is carried out by a broad class of cells known as
hemocytes (Fig 3.) Klowden (2013)(Fig 4.) cites multiple functions of these cells ranging from: phagocytosis (plasmatocytes) and encapsulation (plasmatocytes, oenocytes), to coagulation (granulocytes, oenocytes) and wound healing, as well as various metabolic roles (granulocytes). We currently believe prohemocytes to be stem-cells and the progenitors of the different, more specialized hemocytes. It is not currently known is adipohemocytes and spherules are true hemocytes, as their functions are origins are currently unknown.
Recognition of non-host material in Manduca sexta and Tenebrio molitor
To study the effects of non-host material on the magnitude and type of immune responses insects, we cut cuticle from two standard lab insects, a lepidopteran (Manduca sexta), and a coleopteran (T. molitor)(Fig. 5-12).
 |
| Figure 5. Start here! |
Once the slips were dry, we removed the M. sexta larva from the ice and removed it from its cup. At this point, the larva was very slow and easy to manipulate. We gently snipped the larva's cuticle so that hemolymph would ooze out (Fig. 7). The hemolymph was collected onto a small piece of parafilm. We then used a micropipet to collect the liquid and pipet it onto the coverslips with attached cuticle (Fig. 9).
 |
| Figure 6. Anesthetizing M. sexta. |
The purpose of pipetting the hemolymph onto the cuticle is to allow the hemocytes in M. sexta to respond to cuticle of another individual of M. sexta, as well as an individual of T. molitor.
 |
| Figure 7. Extracting M. sexta hemolymph |
 |
| Figure 8. Combining cuticle and hemolymph. |
After mixing the cuticle and hemolymph together, we went through a series of steps to visualize the outcomes of the cuticle-hemolymph interactions. The method of visualization we used is called immunolabeling. This method takes advantage of antibodies which bind to specific regions, which when looked at under a microscope at specific wavelengths, will glow. Antigens that bind to the tissue of choice and then are able to be visualized are referred to as primary (1°) antibodies. Some antigens require both a primary antibody, and then a secondary (2°) antibody to be visualized. We used two primary antibodies (Peanut Lectin Algluttin (PNA) which binds granular cells (red on images), and MS34 which binds cell adhesion proteins of plasmatocytes (green on images)) and one secondary antibody (FITC, which binds to MS34) in our study. Lastly, we used a DAPI stain to visualize nuclei (blue) in our samples. Labeling was modified from Willott et al. 1994 Eur. J. Cell Biol.
Results and discussion of the study
Our results are in found in Figs. 1 (M. sexta) and 2 (T. molitor). The stains are in the following order, top to bottom: DAPI (blue), FITC (green), and PNA (red). The stains visualize nuclei, plasmatocytes, and granulocytes, respectively. Images in the first column are the
cuticle of respective insect. Thus, images in the second column are of nuclei and hemocytes.
The first result of our study is that it appears that M. sexta has many more nuclei than T. molitor. Four possible explanations exist: the DAPI stain didn't work as well with T. molitor as with M. sexta, the piece of T. molitor cuticle is physically smaller, and thus, less to visualize, the magnification of the T. molitor cuticle may be larger, showing less of a field, making comparison difficult, and it could be that cells in M. sexta are smaller than in T. molitor. I think the most parsimonious answer is probably that the the nuclei are different sizes.
The second and third result of the study is that M. sexta exhibit a much greater response in terms of the number of plasmatocytes and granulocytes created in response to non-host tissue. It's possible that T. molitor have a large response, but it takes more time for the response to occur. It's also possible that the labels did not work perfectly, and that there was uneven spread of the hemolymph, causing uneven (and perhaps even no) binding in certain regions of the cuticle. It's interesting to see the response of M. sexta against another individual of the same species. This reinforces the points discussed above.
Overall, I would say the experiment should be done again to confirm these results, and to reduce out any potential confounding factors discussed here. I hope you learned something. I apologize for the formatting, Blogger sucks apparently.
 |
Figure 9. Pipetting clear liquids   |

References
Klowden, M. J. 2013. Physiological Systems in Insects
3rd ed. Elsevier, San Diego, CA.
Willot et al. 1994. Europ. J. Cell Biol.